what is the medical term that means state of having a fixed relation to a pathogen?
Full general Concepts
Pathogenesis
Pathogenesis is the process past which an infection leads to disease. Pathogenic mechanisms of viral illness include (one) implantation of virus at the portal of entry, (2) local replication, (3) spread to target organs (disease sites), and (iv) spread to sites of shedding of virus into the surroundings. Factors that affect pathogenic mechanisms are (i) accessibility of virus to tissue, (2) cell susceptibility to virus multiplication, and (3) virus susceptibility to host defenses. Natural selection favors the say-so of low-virulence virus strains.
Cellular Pathogenesis
Direct cell harm and death from viral infection may effect from (1) diversion of the cell's energy, (2) shutoff of jail cell macromolecular synthesis, (3) contest of viral mRNA for cellular ribosomes, (4) competition of viral promoters and transcriptional enhancers for cellular transcriptional factors such as RNA polymerases, and inhibition of the interferon defence mechanisms. Indirect jail cell impairment can result from integration of the viral genome, consecration of mutations in the host genome, inflammation, and the host immune response.
Tissue Tropism
Viral analogousness for specific torso tissues (tropism) is determined by (1) cell receptors for virus, (2) cell transcription factors that recognize viral promoters and enhancer sequences, (3) ability of the prison cell to support virus replication, (4) concrete barriers, (five) local temperature, pH, and oxygen tension enzymes and non-specific factors in trunk secretions, and (vi) digestive enzymes and bile in the gastrointestinal tract that may inactivate some viruses.
Implantation at the Portal of Entry
Virions implant onto living cells mainly via the respiratory, gastrointestinal, skin-penetrating, and genital routes although other routes can be used. The concluding upshot of infection may be determined past the dose and location of the virus as well as its infectivity and virulence.
Local Replication and Local Spread
Well-nigh virus types spread amid cells extracellularly, simply some may also spread intracellularly. Institution of local infection may lead to localized disease and localized shedding of virus.
Dissemination from the Portal of Entry
Viremic: The near mutual route of systemic spread from the portal of entry is the apportionment, which the virus reaches via the lymphatics. Virus may enter the target organs from the capillaries by (1) multiplying in endothelial cells or fixed macrophages, (two) diffusing through gaps, and (three) being carried in a migrating leukocyte.
Neural: Broadcasting via nerves usually occurs with rabies virus and sometimes with herpesvirus and poliovirus infections.
Incubation Menses
The incubation menses is the time between exposure to virus and onset of disease. During this unremarkably asymptomatic period, implantation, local multiplication, and spread (for disseminated infections) occur.
Multiplication in Target Organs
Depending on the balance between virus and host defenses, virus multiplication in the target organ may be sufficient to cause affliction and expiry.
Shedding of Virus
Although the respiratory tract, alimentary tract, urogenital tract and blood are the near frequent sites of shedding, various viruses may be shed at virtually every site.
Congenital Infections
Infection of the fetus as a target "organ" is special considering the virus must traverse additional physical barriers, the early on fetal immune and interferon defense systems may exist immature, transfer of the maternal defenses are partially blocked by the placenta, the developing first-trimester fetal organs are vulnerable to infection, and hormonal changes are taking identify.
Introduction
Pathogenesis is the process by which virus infection leads to disease. Pathogenic mechanisms include implantation of the virus at a body site (the portal of entry), replication at that site, then spread to and multiplication inside sites (target organs) where affliction or shedding of virus into the environs occurs. Near viral infections are subclinical, suggesting that body defenses against viruses arrest virtually infections before disease symptoms become manifest. Knowledge of subclinical infections comes from serologic studies showing that sizeable portions of the population have specific antibodies to viruses even though the individuals have no history of disease. These inapparent infections have great epidemiologic importance: they constitute major sources for dissemination of virus through the population, and they confer amnesty (see Ch. 48).
Many factors affect pathogenic mechanisms. An early on determinant is the extent to which trunk tissues and organs are attainable to the virus. Accessibility is influenced past concrete barriers (such as fungus and tissue barriers), by the distance to be traversed inside the body, and by natural defense mechanisms. If the virus reaches an organ, infection occurs only if cells capable of supporting virus replication are present. Cellular susceptibility requires a prison cell surface attachment site (receptor) for the virions and as well an intracellular environs that permits virus replication and release. Fifty-fifty if virus initiates infection in a susceptible organ, replication of sufficient virus to cause disease may be prevented past host defenses (run across Chs. 49 and 50).
Other factors that determine whether infection and affliction occur are the many virulence characteristics of the infecting virus. To cause disease, the infecting virus must be able to overcome the inhibitory furnishings of concrete barriers, distance, host defenses, and differing cellular susceptibilities to infection. The inhibitory effects are genetically controlled and therefore may vary amidst individuals and races. Virulence characteristics enable the virus to initiate infection, spread in the body, and replicate to large enough numbers to impair the target organ. These factors include the power to replicate under sure circumstances during inflammation, during the febrile response, in migratory cells, and in the presence of natural trunk inhibitors and interferon. Extremely virulent strains often occur within virus populations. Occasionally, these strains become dominant as a issue of unusual selective pressures (see Ch. 48). The viral proteins and genes responsible for specific virulence functions are only but beginning to be identified.
Fortunately for the survival of humans and animals (and hence for the infecting virus), almost natural selective pressures favor the dominance of less virulent strains. Because these strains practice non cause severe disease or death, their replication and transmission are not impaired by an incapacitated host. Mild or inapparent infections can result from absence of 1 or more virulence factors. For instance, a virus that has all the virulence characteristics except the ability to multiply at elevated temperatures is arrested at the febrile stage of infection and causes a milder disease than its totally virulent counterpart. Live virus vaccines are composed of viruses scarce in one or more virulence factors; they cause only inapparent infections and yet are able to replicate sufficiently to induce immunity.
The occurrence of spontaneous or induced mutations in viral genetic fabric may alter the pathogenesis of the induced illness, east.k. HIV. These mutations tin can be of particular importance with the development of drug resistant strains of virus.
Disease does non always follow successful virus replication in the target organ. Disease occurs just if the virus replicates sufficiently to damage essential cells directly, to crusade the release of toxic substances from infected tissues, to impairment cellular genes or to impairment organ office indirectly as a consequence of the host immune response to the presence of virus antigens.
As a group, viruses use all conceivable portals of entry, mechanisms of spread, target organs, and sites of excretion. This abundance of possibilities is not surprising considering the astronomic numbers of viruses and their variants (encounter Ch. 43).
Cellular Pathogenesis
Straight jail cell damage and death may result from disruption of cellular macromolecular synthesis by the infecting virus. Also, viruses cannot synthesize their genetic and structural components, and so they rely almost exclusively on the host prison cell for these functions. Their parasitic replication therefore robs the host prison cell of free energy and macromolecular components, severely impairing the host'south power to part and often resulting in cell death and illness.
Pathogenesis at the cellular level can be viewed as a procedure that occurs in progressive stages leading to cellular affliction. As noted in a higher place, an essential aspect of viral pathogenesis at the cellular level is the contest between the synthetic needs of the virus and those of the host cell. Since viruses must use the jail cell's machinery to synthesize their own nucleic acids and proteins, they have evolved various mechanisms to subvert the cell's normal functions to those required for production of viral macromolecules and eventually viral progeny. The function of some of the viral genetic elements associated with virulence may be related to providing conditions in which the synthetic needs of the virus compete effectively for a limited supply of cellular macromolecule components and synthetic machinery, such equally ribosomes.
Damage of cells by replicating virus and damage by the immune response are considered farther in Chapters 44 and 50, respectively.
Tissue Tropism
Most viruses have an affinity for specific tissues; that is, they display tissue specificity or tropism. This specificity is determined by selective susceptibility of cells, physical barriers, local temperature and pH, and host defenses. Many examples of viral tissue tropism are known. Polioviruses selectively infect and destroy certain nerve cells, which have a higher concentration of surface receptors for polioviruses than do virus-resistant cells. Rhinoviruses multiply exclusively in the upper respiratory tract because they are adapted to multiply best at depression temperature and pH and high oxygen tension. Enteroviruses tin can multiply in the intestine, partly because they resist inactivation by digestive enzymes, bile, and acid. The cell receptors for some viruses take been identified. Rabies virus uses the acetylcholine receptor present on neurons equally a receptor, and hepatitis B virus binds to polymerized albumin receptors found on liver cells. Similarly, Epstein-Barr virus uses complement CD21 receptors on B lymphocytes, and human immunodeficiency virus uses the CD4 molecules present on T lymphocytes as specific receptors.
Viral tropism is as well dictated in office by the presence of specific jail cell transcription factors that require enhancer sequences within the viral genome. Recently, enhancer sequences have been shown to participate in the pathogenesis of certain viral infections. Enhancer sequences within the long terminal repeat (LTR) regions of Moloney murine leukemia retrovirus are active in certain host tissues. In improver, JV papovavirus appears to accept an enhancer sequence that is active specifically in oligodendroglia cells, and hepatitis B virus enhancer activity is most active in hepatocytes. Tissue tropism is considered further in Affiliate 44.
Sequence of Virus Spread in the Host
Implantation at Portal of Entry
Viruses are carried to the trunk by all possible routes (air, food, bites, and any contaminated object). Similarly, all possible sites of implantation (all trunk surfaces and internal sites reached by mechanical penetration) may be used. The frequency of implantation is greatest where virus contacts living cells directly (in the respiratory tract, in the alimentary tract, in the genital tract, and subcutaneously). With some viruses, implantation in the fetus may occur at the time of fertilization through infected germ cells, equally well as later in gestation via the placenta, or at birth.
Fifty-fifty at the primeval stage of pathogenesis (implantation), certain variables may influence the concluding effect of the infection. For instance, the dose, infectivity, and virulence of virus implanted and the location of implantation may decide whether the infection will be inapparent (subclinical) or will crusade mild, severe, or lethal disease.
Local Replication and Local Spread
Successful implantation may be followed by local replication and local spread of virus (Fig. 45-1). Virus that replicates within the initially infected cell may spread to adjacent cells extracellularly or intracellularly. Extracellular spread occurs by release of virus into the extracellular fluid and subsequent infection of the next cell. Intracellular spread occurs by fusion of infected cells with adjacent, uninfected cells or past mode of cytoplasmic bridges between cells. Near viruses spread extracellularly, only herpesviruses, paramyxoviruses, and poxviruses may spread through both intracellular and extra cellular routes. Intracellular spread provides virus with a partially protected surround because the antibiotic defence force does not penetrate cell membranes.
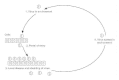
Effigy 45-1
Virus spread during localized infection. Numbers indicate sequence of events.
Spread to cells beyond next cells may occur through the liquid spaces within the local site (eastward.g., lymphatics) or past diffusion through surface fluids such equally the mucous layer of the respiratory tract. As well, infected migratory cells such every bit lymphocytes and macrophages may spread the virus within local tissue.
Institution of infection at the portal of entry may be followed by connected local virus multiplication, leading to localized virus shedding and localized illness. In this manner, local sites of implantation also are target organs and sites of shedding in many infections (Table 45-1). Respiratory tract infections that fall into this category include influenza, the common common cold, and parainfluenza virus infections. Alimentary tract infections acquired by several gastroenteritis viruses (e.chiliad., rotaviruses and picornaviruses) too may fall into this category. Localized pare infections of this blazon include warts, cowpox, and molluscum contagiosum. Localized infections may spread over torso surfaces to infect distant surfaces. An example of this is the picornavirus epidemic conjunctivitis shown in Figure 45-2; in the absence of viremia, virus spreads directly from the eye (site of implantation) to the throat and intestine. Other viruses may spread internally to afar target organs and sites of excretion (disseminated infection). A third category of viruses may cause both local and disseminated disease, as in herpes simplex and measles.
Table 45-1
Pathogenesis of Selected Virus Infection: Localized Infections.

Figure 45-2
Spread of picornavirus over body surfaces from heart to pharynx and intestine during natural infection. Local neutralizing antibiotic action is shown. (Adapted from Langford MP, Stanton GJ, Barber JC: Early appearing antiviral activity in human tears during (more...)
Dissemination from the Portal of Entry
Dissemination in the Bloodstream
At the portal of entry, multiplying virus contacts pathways to the blood and peripheral nerves, the principal routes of widespread dissemination through the body. The most common road of systemic spread of virus involves the apportionment (Fig. 45-3 and Table 45-two). Viruses such as those causing poliomyelitis, smallpox, and measles disseminate through the blood after an initial period of replication at the portal of entry (the comestible and respiratory tracts), where the infection often causes no meaning symptoms or signs of illness because the virus kills cells that are expendable and hands replaced. Virus progeny diffuse through the afferent lymphatics to the lymphoid tissue so through the efferent lymphatics to infect cells in shut contact with the bloodstream (e.k., endothelial cells, specially those of the lymphoreticular organs). This initial spread may effect in a brief primary viremia. Subsequent release of virus directly into the bloodstream induces a secondary viremia, which usually lasts several days and puts the virus in contact with the capillary system of all body tissues. Virus may enter the target organ from the capillaries by replicating within a capillary endothelial cell or stock-still macrophage and then being released on the target organ side of the capillary. Virus may also diffuse through small gaps in the capillary endothelium or penetrate the capillary wall through an infected, migrating leukocyte. The virus may then replicate and spread within the target organ or site of excretion by the same mechanisms as for local broadcasting at the portal of entry. Illness occurs if the virus replicates in a sufficient number of essential cells and destroys them. For instance, in poliomyelitis the central nervous organisation is the target organ, whereas the alimentary tract is both the portal of entry and the site of shedding. In some situations, the target organ and site of shedding may be the same.
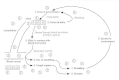
Figure 45-iii
Virus spread through bloodstream during a generalized infection. Numbers indicate sequence of events.
Table 45-2
Pathogenesis of Selected Virus Infections: Disseminated Infections.
Broadcasting in Fretfulness
Broadcasting through the nerves is less mutual than bloodstream dissemination, but is the means of spread in a number of important diseases (Fig. 45-4). This mechanism occurs in rabies virus, herpesvirus, and, occasionally, poliomyelitis virus infections. For instance, rabies virus implanted past a seize with teeth from a rabid fauna replicates subcutaneously and within muscular tissue to attain nervus endings. Evidence indicates that the virus spreads centrally in the neurites (axons and dendrites) and perineural cells, where virus is shielded from antibody. This nerve route leads rabies virus to the central nervous organization, where disease originates. Rabies virus and then spreads centrifugally through the nerves to achieve the salivary glands, the site of shedding. Tabular array 45-2 shows other examples of nervus spread.

Figure 45-iv
Virus spread through nerves during a generalized infection. Numbers indicate sequence of events.
Incubation Period
During nigh virus infections, no signs or symptoms of disease occur through the stage of virus broadcasting. Thus, the incubation period (the time betwixt exposure to virus and onset of affliction) extends from the time of implantation through the phase of dissemination, catastrophe when virus replication in the target organs causes disease. Occasionally, mild fever and malaise occur during viremia, merely they often are transient and have petty diagnostic value.
The incubation period tends to be brief (1 to 3 days) in infections in which virus travels merely a curt altitude to reach the target organ (i.e., in infections in which affliction is due to virus replication at the portal of entry). Conversely, incubation periods in generalized infections are longer because of the stepwise fashion by which the virus moves through the trunk earlier reaching the target organs. Other factors also may influence the incubation menstruation. Generalized infections produced by togaviruses may have an unexpectedly short incubation menstruum because of directly intravascular injection (insect bite) of a rapidly multiplying virus. The mechanisms governing the long incubation catamenia (months to years) of persistent infections are poorly understood. The persistently infected prison cell is often not lysed, or lysis is delayed. In improver, disease may result from a belatedly immune reaction to viral antigen (e.g., arenaviruses in rodents), from unknown mechanisms in ho-hum viral infections during which no allowed response has been detected (as in the scrapie-kuru group), or mutation in the host genetic cloth resulting in cellular transformation and cancer.
Multiplication in Target Organs
Virus replication in the target organ resembles replication at other trunk sites except that (one) the target organ in systemic infections is unremarkably reached late during the stepwise progression of virus through the body, and (2) clinical disease originates there. At each stride of virus progression through the trunk, the local recovery mechanisms (local body defenses, including interferon, local inflammation, and local amnesty) are activated. Thus, when the target organ is infected, the previously infected sites may take reached various stages of recovery. Figure 45-ii illustrates this staging of infection and recovery in different tissues during a spreading surface infection. Circulating interferon and allowed responses probably account for the termination of viremia, but these responses may be also late to prevent seeding of virus into the target organ and into sites of shedding. Nevertheless, these systemic defenses can lengthened in diverse degrees into target organs and thereby help retard virus replication and affliction.
Depending on the balance between virus and host defenses (run into Chs. 49 and fifty), virus multiplication in the target organ may be sufficient to produce dysfunction manifested past disease or death. Additional constitutional affliction such as fever and malaise may event from improvidence of toxic products of virus replication and cell necrosis, as well every bit from release of lymphokines and other inflammatory mediators. Release of leukotriene C4 during respiratory infection may crusade bronchospasm. Viral antigens also may participate in allowed reactions, leading to disease manifestations. In addition, damage of leukocytes and immunosuppression by some viruses may cause secondary bacterial infection.
Shedding of Virus
Considering of the diversity of viruses, almost every possible site of shedding is utilized (Table 45-2); however, the most frequent sites are the respiratory and alimentary tracts. Blood and lymph are sites of shedding for the arboviruses, since biting insects become infected past this route. HIV is shed in blood and semen. Milk is a site of shedding for viruses such as some RNA tumor viruses (retroviruses) and cytomegalovirus (a herpesvirus). Several viruses (e.1000., cytomegaloviruses) are shed simultaneously from the urinary tract and other sites more unremarkably associated with shedding. The genital tract is a common site of shedding for herpesvirus type 2 and may be the road through which the virus is transmitted to sexual partners or the fetus. Saliva is the primary source of shedding for rabies virus. Cytomegalovirus is as well shed from these last 2 sites. Finally, viruses such equally tumor viruses that are integrated into the Dna of host cells tin be shed through germ cells.
Congenital Infections
Infection of the fetus is a special case of infection in a target organ. The factors that determine whether a target organ is infected also apply to the fetus, simply the fetus presents boosted variables. The allowed and interferon systems of the very immature fetus are immature. This immaturity, coupled with the partial placental barrier to transfer of maternal immunity and interferon, deprive the very young fetus of important defence force mechanisms. Some other variable is the high vulnerability to disruption of the apace developing fetal organs, particularly during the kickoff trimester of pregnancy. Furthermore, susceptibility to virus replication may exist modulated by the undifferentiated state of the fetal cells and past hormonal changes during pregnancy. Although virus multiplication in the fetus may pb to congenital anomalies or fetal decease, the mother may take simply a mild or inapparent infection.
To cause built anomalies, virus must attain the fetus and multiply in it, thereby causing maldeveloped organs. Generally, virus reaches the fetus during maternal viremia by infecting or passing through the placenta to the fetal apportionment and so to fetal target organs. Sufficient virus multiplication may disrupt development of fetal organs, particularly during their rapid development (the start trimester of pregnancy). Although many viruses occasionally crusade built anomalies, cytomegalovirus and rubella virus are the nearly mutual offenders. Virus shedding past the congenitally infected newborn infant may occur equally a result of persistence of the virus infection at sites of shedding.
References
-
Albrecht T, Boldogh I, Fons M. et al. Cell activation signals and the pathogenesis of human cytomegalovirus. Intervirology. 1990;31:68. [PubMed: 2165048]
-
Coen DM. Acyclovir-resistant, pathogenic herpesviruses. Trends Microbiol. 1994;ii:481. [PubMed: 7889324]
-
Fields BN. How do viruses crusade different diseases? J Am Med Assoc. 1983;250:1754. [PubMed: 6310171]
-
Grieder FB, Davis NL, Aronson JF. et al. Specific restrictions in the progression of Venezuelan equine encephalitis virus-induced disease resulting from single amino acrid changes in glycoproteins. Virology. 1995;206:994. [PubMed: 7856110]
-
Singh IP, Chopra AK, Coppenhaver DH. et al. Vertebrate brains contain a broadly active antiviral substance. Antiviral Inquiry. 1995;27:375. [PubMed: 8540757]
-
Strayer DS, Laybourne KA, Heard HK. Determinants of the power of malignant fibroma virus to induce immune dysfunction and tumor dissemination in vivo. Microb Pathos. 1990;9:173. [PubMed: 1964998]
-
Wold We, Hermiston TW, Tollefson AE. Adenovirus proteins that subvert host defenses. Trends Microbiol. 1994;ii:437. [PubMed: 7532531]
Source: https://www.ncbi.nlm.nih.gov/books/NBK8149/
0 Response to "what is the medical term that means state of having a fixed relation to a pathogen?"
Post a Comment